|
| Eng |
Управление популяцией саранчовых в степных ландшафтах. Современные
подходы и технологии
| ||||||||||||||||||||||||||||||||||||||||||||||||
|
Район |
Заселено саранчовыми, |
Обработано, |
|
Нижнее Поволжье - Северный Кавказ |
887 (1,3) |
Около 350 (0,5) |
|
Европейские лесостепи |
58 (0,2) |
1 (менее 0,1) |
|
Южный Урал |
1460 (4,3) |
Около 780 (2,3) |
|
Юг Западной Сибири |
840-1100 (0,9-1,2) |
Около 1600 (1,7) |
|
Юг Средней и Восточной Сибири |
250 (0,1) |
150 (менее 0,1) |
При оценке этих данных нужно помнить, что некоторые участки обрабатывались неоднократно.
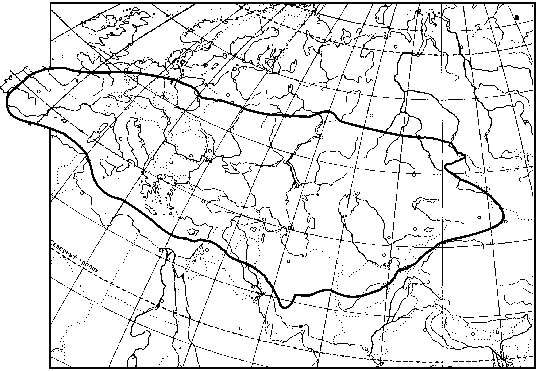
На исходе XX века основной район массового размножения итальянского пруса сместился на Южный Урал, в Кулунду и Прииртышье. Это классическаяобласть его массовых размножений. Здесь они отмечались в 1922, 1938, 1943-1945, 1952-1956, 1967-1971 и 1977-1982 гг., причем местами плотность пруса превышала 2000 особей на м2 [Архангельский, 1957; Александров, 1962 и др.].
Как известно, в Кулундинской степи и в Прииртышье наиболее благоприятные условия для нарастания численности итальянского пруса возникают после нескольких лет с теплой и засушливой весной. Связано это в первую очередь со снижением эмбриональной смертности [ср. Сергеев и др., 1995].
Именно благоприятные погодные условия были одной из основных причин того, что лето 1999 г. оказалось на юго-востоке Западной Сибири и в сопредельном Казахстане необычайно сложным. Май был сухим и теплым. Июнь был также сравнительно теплым, но с умеренным количеством осадков, что способствовало интенсивному росту трав. В результате резкий и во многом неожиданный подъем численности саранчовых захватил почти все степи этого региона. Этому благоприятствовали и уже отмечавшиеся социально-экономические и политические перемены последнего десятилетия. Ситуация усугублялась тем, что прилегающие степные районы Казахстана также были охвачены вспышкой итальянского пруса. Местные условия были еще более благоприятными для размножения и развития этого вида. В значительной степени это определялось и определяется гораздо большими площадями залежей. Как известно, во многих районах Казахстана заброшено до 50 и более процентов ранее ежегодно распахивавшихся массивов. С территории Казахстана стаи взрослых саранчовых могли залетать и на территорию России. Естественно, могли происходить и обратные перелеты.
Высокая плотность пруса была наиболее типична для его любимых местообитаний - это степные участки и особенно молодые залежи с большим количеством разнотравья и легкими песчаными или песчанистыми почвами. Именно такие залежи, а также придорожные полосы и лесополосы стали основными участками, где происходил рост численности. Уже отсюда саранчовые при недостатке корма могли перемещаться в другие места, в том числе поля. Именно в это время (когда личинки старших возрастов и взрослые начали мигрировать, в том числе расселяться на поля) подъем численности и привлек к себе внимание. Соответственно меры по управлению численностью оказались не вполне адекватными.
Сказалась нерегулярность вспышек, особенно в этих районах, и отсутствие достаточного финансирования в межвспышечный период как для организации исследований и мониторинга, так и для накопления резервов. Особенно остро, кроме запоздалой реакции, проявилось отсутствие необходимых технических возможностей, а также препаратов, оптимальных для снижения численности. Так, очень часто использовались cинтетические пиретроиды.
По нашим данным, летом 1999 г. популяции итальянского пруса в Кулунде занимали промежуточное положение между типично стадной и нестадной формами. Можно предполагать, что во второй половине лета в Кулундинской степи сформировались смешаные популяции, частично местного происхождения, а частично сложившиеся из особей, прилетевших из Казахстана. Во многих местах, благоприятных для яйцекладки, была обнаружена высокая плотность кубышек. Если учесть, что каждая кубышка может содержать 20-50 яиц, то в случае благоприятных погодных условий следовало ожидать дальнейшего нарастания численности в 2000 году.
В результате в 2000 г., несмотря на прохладный и дождливый май и связанную с этим высокую эмбриональную смертность, плотность пруса в Кулунде была в среднем существенно выше, чем в 1999 г. Вместе с тем развитие пруса проходило со значительным опозданием - примерно на 10 дней по сравнению со средним многолетним. Вскоре после массового отрождения, т.е. во время преобладания личинок 1-го возраста, средняя плотность достигала 11-25 особей на квадратный метр, в кулигах же она нередко превышала 400-1000 особей на такую же площадь (20 июня). Кулиги личинок младших возрастов в основном держались в местах отрождения, лишь немного смещаясь ближе к сочному разнотравью и на края полей кормовых культур. Первые личинки 4-го возраста появились 25 июня. 5-6 июля были многочисленны личинки 4-5-го возрастов. Первый взрослый самец пруса был обнаружен 7 июля. Подобная динамика вполне соответствует 10-дневному сдвигу. Кулиги личинок средних и старших возрастов перемещались более активно и заселяли в первую очередь участки с богатыми кормовыми ресурсами. Нередко перемещения шли вдоль полевых дорог.
Обследование в второй половине июля показало, что в южной части Кулунды средняя плотность пруса существенно уменьшилась по сравнению с 1999 г., тогда как на обширной территории от Михайловского района Алтайского края до Карасукского района Новосибирской области она, наоборот, увеличилась в 2-13 раз. Естественно, при такой плотности взрослой саранчи отложено значительное число кубышек. Обследованные популяции, так же как и в предыдущем году, занимали промежуточное положение между стадной и одиночной формами. Все это означало, что с крайне большой вероятностью вспышка будет развиваться дальше и в некоторых районах, например, на северо-западе Алтайского края, можно ожидать следующего витка нарастания численности. Также можно ожидать дальнейшего подъема численности в других степных и лесостепных регионах России и Казахстана.
Этот прогноз в целом подтвердился. В начале лета 2001 г. высокая и средняя численность пруса, а также нестадных саранчовых, наблюдалась на значительных территориях юга Западной Сибири, а на севере Дагестана и Ставропольского края обозначился новый очаг вспышки массового размножения пруса.
Это означает, что по-прежнему актуальны проблемы, связанные с охраной природы и поддержанием биологического разнообразия, так как, несомненно, значительные участки будут перепахиваться, а затем будут использоваться инсектициды. Как известно, и то и другое приводит к значительному сокращению разнообразия живых организмов на конкретных участках. Кроме того, использование для снижения численности саранчовых такого метода, как распашка залежей и неудобиц, на самом деле крайне рискованно, поскольку после распашки (если что-то не посеять) снова возникает молодая залежь, еще более подходящая для пруса. Реально в таких случаях необходима не распашка, а ускорение восстановления степной экосистемы. В любом случае дальнейший подъем численности саранчовых потребует значительных финансовых вливаний как со стороны федеральных, так и местных властей. Кроме того, по-прежнему актуальной будет проблема взаимоотношений не только между регионами, но и между соседними государствами.
Богатейший мировой опыт управления численностью саранчовых в той области, которую обычно называют "борьба", показывает, что наблюдаемая в природе картина очень сложна, а наши представления до сих пор остаются во многом далекими от действительности. Как правило, нарастание численности саранчовых обнаруживают с запозданием, бросают на “подавление” вспышки подручные средства (естественно, далеко не лучшие) и надеются, что крупномасштабные обработки инсектицидами помогут спасти урожай. Реально необходимы длительные наблюдения, установление закономерностей развития популяций саранчовых и соответствующее изменение стратегических установок с “тушения пожара” на проведение превентивных мероприятий (таких, например, как уменьшение площади молодых залежей).
Характер это сложности задается, по крайней мере, тремя основными компонентами:
(1) Географическая и временная неоднородность среды обитания саранчовых, выражающаяся в первую очередь в том, что каждый район, каждый участок не похож на другой. Более того, состояние любого участка, включая столь важные для жизни саранчовых показатели, как уровень увлажнения и теплообеспеченности, особенности почв и растительного покрова, постоянно меняется. Весьма своеобразен в каждом конкретном случае и характер влияния человека.
(2) Различия видов, сложившиеся в ходе их возникновения. Хорошо известно, что даже близкие виды не похожи друг на друга. Эти различия могут проявлятся очень резко, но нередко прослеживаются только при тщательном исследовании.
(3) Различия между разными популяциями каждого вида связаны с их наследуемыми особенностями и обычно менее заметны. Но принципиальные различия между популяциями одного вида в разных районах, допустим, у тех же стадных саранчовых - пустынной и перелетной саранчи, известны давно. Существенно, что даже близлежащие местные популяции могут различаться по экологическим особенностям. Это особенно важно для нестадных саранчовых, так как у большинства из них, судя по имеющимся данным, перемещения между соседними поселениями минимальны.
Стадные и нестадные саранчовые. Чем же различаются подходы к управлению численностью стадных и нестадных саранчовых? Конечно, на стаи саранчовых человек обращал внимание на протяжении многих веков. Но то, что есть виды, образующие как стадную, так и одиночную формы, видимо, первым установил Ф.П. Кеппен в 1870 г. Основы современных представлений о так называемом фазовом полиморфизме (т.е. о возможном наличии у одного и того же вида двух разных форм, различающихся по морфологии, физиологии и поведению) были заложены Б.П. Уваровым в 20-е годы XX века [Uvarov, 1977 и др.]. С этого времени и сохраняется некоторая неопределенность в употреблении слов стадный и нестадный.
Мы будем рассматривать термины стадный, нестадный и переходный как видовые характеристики (даже признаки), сложившиеся в ходе эволюционного освоения той или иной экологической ниши. Сейчас можно говорить о том, что стадные, нестадные и переходные саранчовые различаются по следующим основным признакам (таблица 2).
Таблица 2. Основные различия стадных, нестадных и переходных видов саранчовых
|
Стадные виды |
Переходные виды |
Нестадные виды |
|
Морфологические особенности |
||
|
Явные различия особей, живущих по одиночке, и особей, живущих в группах. Эти различия могут проявляться в относительной длине крыльев, конечностей, цветовых вариациях, форме переднеспинки |
Могут быть некоторые различия, часто чисто статистические |
Нет различий |
|
Эколого-поведенческие особенности |
||
|
Образование кулиг и(или) стай, каждая из которых объединяет саранчовых, движущихся практически однонаправленно |
Возможно образование небольших кулиг и(или) стай |
Образование только скоплений с повышенной численностью, часто в связи с яйцекладкой |
|
Характер миграций |
||
|
Способны к дальним миграциям (свыше нескольких сотен километров) |
Способны к ограниченным миграциям (порядка десятков и сотен километров) |
Обычны только внутрипопуляционные перемещения |
|
Примеры |
||
|
Пустынная саранча, перелетная саранча |
Итальянский прус, сенегальская кобылка, мароккская саранча |
Белополосая, крестовая, темнокрылая кобылки |
Несомненно, четких границ между этими группами саранчовых нет. Давно известно, что есть типичные стадные виды, такие как пустынная и перелетная саранча, а есть и типичные "одиночники" - та же белополосая кобылка. Вместе с тем итальянская и мароккская саранча ближе к классическим стадным видам, а сибирская кобылка, хотя и проявляет в годы подъемов численности элементы стадности, тем не менее, все-таки принадлежит к нестадными саранчовыми.
На протяжении десятилетий основной упор в управлении численностью делался на стадных саранчовых. Такое внимание совершенно оправдано. Вместе с тем для России и большинства друг стран СНГ, а также США и Канады, актуальна проблема нестадных саранчовых, ущерб от которых может быть весьма значительным.
Именно поэтому необходимо попытаться определить различия в подходах к управлению численностью стадных и нестадных видов. Насколько реально использование богатого опыта борьбы со стадными видами? И, напротив, можно ли применять результаты, полученные для нестадных видов, для стадных саранчовых?
Наиболее важны следующие различия:
(1) Управление численностью нестадных форм, как правило, направлено на все многовидовое сообщество. Хотя часто один из видов является господствующим (например, доля сибирской или белополосой кобылки в сообществе может составлять 80-90%), тем не менее, обычно подъем численности хорошо прослеживается у двух и более видов.
(2) Многовидовые сообщества нестадных саранчовых часто характеризуются тем, что высокий суммарный уровень численности в них может сохранятся на протяжении всего сезона и даже ряда лет, но входящие в них массовые виды как бы сменяют друг друга во времени.
(3) Нестадные виды, обитающие в одной местности, различаются по многим признакам: размеру, экологическим особенностям (в том числе предпочитаемым местообитаниям и сезонной динамике), пищевым пристрастиям, устойчивости и восприимчивости к инсектицидам.
Следует отметить, что исторически наблюдается изменение экологического и географического распределения многих нестадных и, отчасти, стадных видов. Фактически почти исчезли европейские поселения среднерусской перелетной саранчи, хотя, по данным Алейниковой [1950], еще в 1947-48 гг. она была крайне обильной на юге Татарстана.
Во время вспышек нестадных саранчовых уровень численности может быть значительным. Так, плотность белополосой кобылки в Центральной Якутии превышает 1000 особей/м2. На юге Сибири может быть велика плотность темнополосой кобылки - до 30 экз./м2. Численность сибирской кобылки достигает 200 особей/м2 в Тянь-Шане, до 14-30 особей - на горных пастбищах Южного Алтая и Альп и на перевыбитых пастбищах Центральной Якутии [Сергеев и др., 1995]. При таких уровнях плотности нестадных саранчовые в целом могут уничтожать почти все зеленые части трав.
Многолетнее слежение за состоянием поселений, как итальянского пруса, так и саранчовых вообще - основа любых прогнозов, особенно многолетних и долгосрочных. Чем тщательнее собираются данные во время такого слежения, тем точнее может быть прогноз, тем реже будут чрезвычайные ситуации и тем ниже будут экономические и экологические потери. Следует подчеркнуть, что по некоторым оценкам [PRIFAS, 1989], годовые затраты на защиту полевых культур от саранчовых во время вспышки достаточны для покрытия расходов на предупреждающий контроль на протяжении не менее 15-20 лет!
(1) Наземное слежение. В первую очередь необходимо соблюдение статистических требований. Нужно наличие контроля и отбор достаточного со статистической точки зрения числа проб, так как численность обычно сильно варьирует. В большинстве случаев достаточно 25 таких проб, но нередко (при малой изменчивости) можно ограничиться меньшим их числом. При этом необходимо оценить разброс значений и рассчитать так называемую ошибку выборочной средней (использовав для этого обычный учебник по математической статистике или биометрии).
Выборки должны размещаться на сравнительно однородной по своим природным особенностям территории и должны быть получены в относительно короткие промежутки времени (за время отбора проб погодные условия должны быть неизменными).
При отборе проб нужно избегать соблазна их преимущественного размещения в местах повышенной плотности саранчовых. Необходимо либо размещать участки отбора проб по регулярной схеме: например, через равные промежутки вдоль двух перпендикулярных линий, полностью пересекающих обследуемый участок - поле, пастбище, залежь, либо же распределять их случайно.
Обычно для получения полной картины на протяжении календарного года считается достаточным проведение следующих обследований [Лепешкин, 1934б, в; Волков, 1947; Цыпленков, 1970]:
А) весеннее определение плотности кубышек, в том числе в местах их скоплений, и оценка их состояния;
Б) разведка мест отрождения личинок;
В) наблюдения за перелетами стай взрослых и обследование территорий с низкой плотностью;
Г) осеннее определение плотности кубышек и оценка их состояния.
При каждом обследовании фиксируются место и условия конкретного местообитания, время его проведения, желательно получение данных о микроклимате (есть современные устройства на микропроцессорах, которые позволяют это сделать).
Оценка эффективности обработок. В первую очередь необходимо получение оценок плотности как итальянского пруса в целом, так и по отдельным возрастам личинок и имаго. Необходимо наличие контрольных участков. Во всех случаях целесообразно проводить учеты до обработки и через одни сутки после нее. Желательно и проведение еще нескольких учетов с интервалом в 1-3 дня в зависимости от продолжительности действия препарата. Это позволяет получить данные по развитию популяции.
Обычно для определения эффективности мероприятий по управлению численности используют следующий способ [Семевский, 1968 и др.]:
Э=100(1-КдОп/КпОд),
где Э - биологическая эффективность, %; Кд - плотность в контроле до обработки, Кп - после обработки; Од - в опыте до обработки, Оп - после обработки.
Оценка величины изъятой массы растений и потерь урожая. Для определения масштаба повреждений, наносимых саранчовыми, можно использовать разные методы.
А) Опытные садки. В зависимости от целей и условий может варьировать количество саранчовых и их возраст. Каждый садок должен быть открыт снизу. Все садки должны быть одинаковых размеров. На полях целесообразно, чтобы каждый садок включал равное число растений. На пастбищах и залежах характер растительности внутри садков должен быть сравнительно однородным. Число садков в каждом варианте (например, с 5 личинками 3-го возраста) должно быть достаточным для статистических оценок. Это же относится и к обязательным контрольным садкам, вообще не содержащим саранчовых. При длительных наблюдениях необходимо удалять мертвых насекомых и заменять их на соответствующие живые экземпляры [Wewetzer et al., 1993]. В конце эксперимента на пастбищах и залежах удаляется, высушивается и взвешивается все надземные части растений, на полях зерновых также определяется урожай зерна. Подобные эксперименты позволяют оценить потери надземной фитомассы или урожая при разной плотности саранчовых.
Б) Удаление саранчовых на каком-то участке, например, с помощью инсектицидов, последующее размещение садков на обработанном и необработанном (т.е. с естественной плотностью саранчовых) участков. В результате можно получить оценку потерь урожая при той или иной численности насекомых [Wewetzer et al., 1993].
В) Размещение на участке, занятом саранчовыми, постоянных трансект (линии или полос) с последующим обследованием площадок через каждые 20 м [см. Wewetzer et al., 1993]. Площадки должны быть одинаковых размеров - в зависимости от плотности насекомых от 0,1 до 1 м2. При обследовании каждого участка определяется плотность саранчовых и степень повреждений с использованием следующей шкалы (адаптированной для наших условий):
0 -- нет повреждений;
1 -- слабые повреждения (10%) -- значительная часть листьев с небольшими повреждениями;
2 -- умеренные повреждения (30%) -- большинство листьев с умеренным повреждением;
3 -- значительные повреждения (60%) -- большая часть листьев сильно повреждена;
4 -- крайне серьезные повреждения (80-100%) -- большая часть листьев полностью съедена;
5 -- полное уничтожение - растения съедены до земли.
Необходимо отмечать тип культуры, стадию ее развития, общее состояние растений, а также повреждения, не связанные с саранчовыми. Первое обследование выполняется сразу после появления саранчи, второе - через две недели. Через 7 недель целесообразно исследование ответа растений на повреждение: проявление компенсаторного роста - угнетение и - отсутствие реакции. При сборе урожая нужно оценить реальный урожай - на статистически значимом числе случайно выбранных площадок по 1 м2 срезаются все культурные растения и взвешиваются все зерна.
Для определения потерь можно использовать несколько подходов (предполагая, правда, сходство условий) [Wewetzer et al., 1993]:
(2) Возможности использования персональных компьютеров. В последние годы создано несколько компьютерных программных комплексов для анализа данных по распределению саранчовых. Большая их часть ориентирована на пустынную саранчу и вряд ли может быть адаптирована для саранчовых степных районов Евразии.
Однако в США была разработана довольно простая, но эффективная экспертная система "Прыгунчик", рассчитанная на использование фермерами. После введения в компьютер необходимой информации (погодные условия, почвы, общая численность саранчовых, экономические параметры) владелец земли сам может оценить необходимость проведения защитных мероприятий и принять то или иное решение. Позднее в Университете Вайоминга была создана более совершенная экспертная система, работающая только в пределах этого штата. С ее помощью владелец земли может получить ответы на важные для него вопросы: насколько серьезна проблема, есть ли конкуренция за корм между саранчовыми и домашними животными.
Разработка подобных программных комплексов для России - дело будущего, будем надеяться близкого. Конечно, в России обеспеченность сельских хозяйств компьютерами оставляет желать много лучшего, но развивать такие подходы необходимо, в том числе и потому, что с развитием хозяйственной самостоятельности сельских производителей затраты на проведение защитных мероприятий им придется оплачивать из своего кошелька. Кроме того, они будут вынуждены оценивать “экологическую чистоту” своей продукции.
(3) Дистанционное слежение и перспективы использования географических информационных систем. Наземное слежение за численностью саранчовых крайне трудоемко. Кроме того, при большом объеме данных очень сложен и их анализ. Именно поэтому в последние годы при определении стратегии и тактики мероприятий по управлению численностью саранчовых активно используются данные спутниковых съемок поверхности Земли. Современные технологии позволяют различать на снимках все разнообразие экосистем, в том числе нарушенных. Например, для пустынной саранчи просто и быстро выявляются островки зеленой растительности в пустынях, что позволяет определять пути перемещения ее стай.
Данные космических снимков можно использовать только тогда, когда есть набор участков, хорошо изученных традиционными методами. Но этого мало, такие участки нужно сопоставить со снимками и соответственно определить их географические координаты. Последнее возможно при использовании топографических карт и устройств определения глобального местонахождения (GPS), позволяющих по сигналам со спутников очень точно определить положение на поверхности Земли.
После сопоставления наземных наблюдений и расшифровок космических снимков на последних можно выделить участки, благоприятные для подъемов численности. Эти данные могут быть внесены в географические информационные системы, использующие компьютерные карты. Обычно такие системы основаны на использовании подобных карт, представленных в виде слоев, каждый из которых содержит ту или иную информацию. Изменение сочетаний слоев, расчет различных показателей позволяет во многом по-новому взглянуть на реальность, в том числе дает возможность выявить многолетние изменения состояния экосистем и населения саранчовых.
К сожалению, как сами космические снимки, так и компьютеры необходимой производительности, а также соответствующее программное обеспечение, пока очень дороги.
В слежении за нестадными саранчовыми принципиально важны:
(1) Необходимость не менее 3-4 обследований (в том числе двух - кубышек) на протяжении теплого сезона и получение серий конкретных значений не только для всего сообщества, но и для отдельных видов, по крайней мере, массовых.
Это заставляет, во-первых, вспомнить о проблеме пороговых значений численности (экономических порогов вредоносности), при превышении которых считается необходимым проведение мероприятий по снижению численности. Судя по всему, обычно используемые оценки -- около 10 особей/м2 - занижены. В большинстве случаев, особенно на пастбищах и сенокосах, реальный порог составляет 20-30 особей/м2 [Лачининский, Локвуд, Сергеев, 1999 и др.]. Вместе с тем в зависимости от конкретных условий (района, местообитания, времени) этот пороговый показатель может быть очень разным, вероятно, от 1 до 80-100 особей/м2.
Во-вторых, встает проблема точного определения видов на всех стадиях развития - от кубышек до взрослых. В современных условиях такое определение фактически невозможно. А это делает значительную часть наших усилий напрасной. Именно поэтому необходима разработка доступных определителей саранчовых, может быть даже для относительно небольших регионов.
(2) Необходимость покрытия при учетах больших территорий и обследования всего разнообразия ландшафтов. Все это делает обследование трудоемким и сложным. Вместе с тем уже существующий опыт использования географических информационных систем (ГИС) и спутниковых съемок показывает, что обследования можно сделать более избирательными.
(3) Насущная потребность в установлении границ между местными поселениями каждого вида и отдельное рассмотрение каждого из них. Этот подход в настоящее время может быть реализован только с использованием ГИС, аэро- и космосъемок. В действительности, решение этой проблемы на местности крайне трудозатратно и дорого.
(4) Относительная легкость прогнозирования, по крайней мере, некоторых из вспышек нестадных форм на уровне всего сообщества. Во многих районах возможность значительного подъема численности нестадных видов четко связана с засухами. Так, на юге Сибири вспышки, как правило, начинаются в засушливый год с сухой весной, которому предшествуют 1-2 также засушливых года.
Традиционные подходы и технологии управления популяциями. История взаимоотношений человека и саранчовых сложна и многообразна. Для человека это обычно означало и означает в первую очередь поиск совокупности таких подходов, которые позволяли бы эффективно снижать численность саранчовых в случае возможного вреда с их стороны. Конечно, эти подходы менялись на протяжении исторического времени. Во многом эти изменения были естественными и определялись развитием соответствующих технологий и сменой научных представлений о саранчовых и их роли в экосистемах.
Для снижения численности саранчовых использовались всевозможные методы - от простого вылавливания с помощью рук или каких-то иных приспособлений до самолетов и вертолетов, оборудованных по последнему слову техники, мотодельтапланов и аэрозольных генераторов.
Попытки управления популяциями предпринимались и предпринимаются на всех стадиях развития яйца, личинок, взрослых. В целом можно согласиться с В.Н. Зряковским [1927], что все-таки наилучшие результаты обычно достигаются в ходе мероприятий, направленных на личинок. Вместе с тем при оценке различных подходов и технологий нужно помнить не только о их экономической и технической эффективности, но и о их приемлемости с экологической (точнее природоохранной) точки зрения.
В общем виде традиционные подходы можно разделить на три большие группы по типу воздействия: (1) физические, связанные с непосредственным физическим уничтожением саранчовых либо самим человеком, либо с помощью каких-то технических приспособлений; (2) химические, при использовании которых саранчовые гибнут в результате воздействия какого-то химического соединения или комплекса таких соединений; (3) биологические, когда снижение численности достигается за счет использования других живых организмов. Отдельно могут рассматриваться агротехнические методы, связанные с рациональным распределением культур и изменением характера землепользования. Подобные методы нередко весьма эффективны, особенно в отношении итальянского пруса [Лепешкин, 1934а; Бунин, 1979]. Естественно, есть методы, которые можно отнести к разным классам: так, вспашка залежей, с одной стороны, типичный агротехнический метод, а с другой, основной эффект при этом достигается благодаря механическому уничтожению кубышек.
Но все существующие методы можно разделить и по характеру охватываемых территорий: (а) сплошные обработки, при которых пытаются захватить сплошное сравнительно большое пространство; (б) барьерно-блочные обработки, когда, например, тот или иной препарат используется на одних участках, а другие (сравнимые по площади) остаются необработанными, и (в) точечные обработки, в случае которых первоначальное воздействие осуществляется только на небольших участках, не сравнимых по площади со всей территорией.
(1) Физические подходы. Эта группа методов основана на физическом (как правило, просто механическом) сборе и(или) уничтожении саранчовых, поэтому их нередко еще называют механическими. Часто они очень эффективны, но требуют значительных трудовых и временных затрат [Daxl et al., 1994].
К таким методам относятся ручной сбор вредителей, использование для удаления насекомых сачков, ловушек, в том числе канавных, загородок и их всевозможных сочетаний.
Еще в первой половине XX века широко использовалась ручная выборка кубышек. Личинок давили катками, волокушами, скотом. Насекомые отлавливались специальными ловушками на конной или ручной тяге, сетями, мешками и т.п. Для отпугивания летящих стай жгли солому, сено и даже посевы, шумели, используя самые разнообразные приемы вплоть до стрельбы из пушек. Большая часть таких методов не эффективна, а их реализация требует значительных трудозатрат.
Ловушки. Существенно эффективнее сооружение ловчих загородок, канав, их аналогов, в том числе в сочетании друг с другом. Естественно, это методы, пригодные для перемещающихся кулиг [Зряковский, 1927]. При этом необходимо учитывать следующее:
(1) ловушки должны быть размещены так, чтобы они располагались на пути движущейся кулиги; их сооружение должно быть закончено не менее чем за полчаса до ее приближения;
(2) длина канавы и(или) заграждения должна превышать ширину кулиги; оптимальная ширина самой канавы - около 70-80, а глубина - около 90-100 см; на дне канавы желательно разместить более глубокие ямы; все стенки должны быть гладкими и отвесными, а лучше с обратным уклоном или с дополнительным горизонтальным ограждением.
Персонал должен контролировать перемещение кулиги и не допускать ее уход в сторону. Для этого, в частности, можно применять переносные щиты из подручного материала. После сосредоточения кулиги или большей ее части в ловушке, последнюю просто засыпают землей и утрамбовывают.
Распашка залежей и неудобиц. Это традиционный и иногда вполне оправданный подход. Для этого используют как плужную вспашку, так и боронование, а также их сочетание. Именно последнее рекомендуют А.Г. Махоткин с соавторами [1999]: осуществлять вспашку с одновременным использованием тяжелой зубовой бороны. Эти же авторы отмечают и высокую эффективность игольчатой бороны после двух ее проходов.
Вместе с тем подобный подход не является эффективным с экологической точки зрения, так как при распашке останавливается восстановительная сукцессия и процесс восстановления начинается снова. При этом создаются условия, благоприятные для размножения саранчовых (особенно пруса) на следующий год.
Кроме механических методов, возможно (часто только теоретически) использование света (в том числе световых ловушек; правда, как известно, итальянский прус почти не летит на свет), акустических, температурных и электромагнитных воздействий, радиации. Некоторые из таких подходов, по меньшей мере, сомнительны с экологической точки зрения. Особенно это относится к источникам радиации. Другие (электромагнитное излучение) требуют более тщательной оценки.
(2) Химические методы становятся доминирующими с начала XX века. При этом обычно используется либо распространение препарата на каком-то участке тем или иным способом (разбрасывание, распыление, разбрызгивание), либо в каких-то точках размещаются отравленные приманки.
Естественно, состав инсектицидов на протяжении века значительно изменился. Если в начале века применяли главным образом соединения мышьяка, затем такие опасные вещества, как ДДТ и ГХЦГ, еще позже фосфорорганические соединения, то теперь среди препаратов, разрешенных к использованию против саранчовых господствуют синтетические пиретроиды: циперметрин и его производные, дельтаметрин, лямбдацигалотрин, фенвалерат, эсфенвалерат и другие. Еще достаточно хорошо представлена фосфорорганика (малатион, фенитротион, хлорпирифос). Некоторые препараты представляют собой комбинацию инсектицидов разных групп.
Вместе с тем в последние годы появились новые препараты, применение которых, вероятно, позволяет, с одной стороны, достичь существенно более высокого эффекта, а с другой, намного снизить нагрузку на экосистемы.
Следует отметить, что существует специальный Экспертный совет по пестицидам при Организации по сельскому хозяйству и продовольствию ООН. Задача Совета - разработка рекомендаций, касающихся эффективности и безопасности препаратов. Недавно на основе отчетов Экспертного совета А.В. Лачининский [2000] проанализировал, в том числе с экологической точки зрения, все основные современные препараты либо их группы.
Недостатки уже давно применяющихся фосфорорганических соединений (карбофос, дурсбан) следующие - необходимость внесения очень больших доз (обычно значительно больше 200 г действующего вещества на гектар), соответственно очень серьезное загрязнение экосистем и относительно небольшой срок действия - всего несколько суток. При использовании таких препаратов значительное падение численности наблюдается примерно на 2-е сутки. Фосфорорганика очень опасна или среднеопасна для других членистоногих (включая водных) и сравнительно малоопасна для позвоночных. По оценкам Всемирной организации здравохранения (ВОЗ), дурсбан умеренно опасен для человека, а карбофос мало опасен.
Карбаматы (например, бендиокарб) близки по своим характеристикам к органофосфатам.
Разнообразные и широко используемые синтетические пиретроиды (фастак, кинмикс, децис, фьюри, каратэ, шерпа и многие другие) применяются при очень низких дозах (примерно 10-20 г действующего вещества на гектар), но при необходимости снижения численности личинок старших возрастов и взрослых количество вносимого препарата приходится увеличивать, иногда значительно. Кроме того, одно из достоинств пиретроидов, а именно быстрое разложение, при их использовании против саранчовых, особенно итальянского пруса, становится недостатком, так как обычно при высоких уровнях численности каждый участок приходится обрабатывать неоднократно. Вместе с тем пиретроиды действуют очень быстро (так называемый эффект нокдауна), и их эффект часто очевиден уже через несколько часов после обработки. Однако часто это только видимость, при обработке против личинок старшего возраста и взрослых часто наблюдается их оживание. Пиретроиды обычно опасны для всех членистоногих (особенно водных) и почти не опасны для позвоночных (в том числе человека). Однако ВОЗ рассматривает каратэ (действующее вещество - лямбдацигалотрин) как умеренно опасный для человека препарат. Кроме того, некоторые пиретроиды (особенно децис - дельтаметрин) отличаются довольно высокой токсичностью для млекопитающих [Ritchie, Dobson, 1995].
Ингибиторы синтеза хитина (димилин, альсистин, номолт) нарушают формирование покровов и других скелетных элементов саранчовых (и других членистоногих), соответственно их развитие прекращается. Именно это определяет медленность воздействия подобных соединений - максимальный эффект прослеживается только через несколько недель после их внесения. Но защитные свойства проявляются на протяжении нескольких недель, а по некоторым данным, проявляются даже на следующий год. Рекомендуемые дозы таких препаратов невелики и составляют от 25 до 60 г действующего вещества на гектар, однако использовать их можно только тогда, когда большая часть саранчовых представлена личинками младших и средних возрастов. Ингибиторы синтеза хитина весьма опасны для всех личинок членистоногих (особенно водных), но практически безопасны для взрослых домашних пчел и, естественно, позвоночных животных.
Преимущество сравнительно нового препарата адонис из группы фенилпиразолов заключается в его хорошо выраженном избирательном воздействии на саранчовых. Поэтому его рекомендуется применять в сверхнизких дозировках (1-5 г действующего вещества на гектар). Наибольшее снижение численности саранчовых обычно наблюдается через 2-3 дня после обработки. Другое свойство адониса - длительное (до нескольких недель) воздействие на саранчовых - одновременно его достоинство и недостаток, так как, с одной стороны, избавляет от необходимости повторных обработок и дает возможность применения отравленных приманок, а с другой, надолго (примерно на месяц - не менее чем на 21 день) выводит обработанный участок из оборота. По существующим оценкам, адонис может представлять опасность для всех наземных членистоногих, но при рекомендованных дозировках практически безопасен для позвоночных и для обитателей водных экосистем.
Кроме того, недавно зарегистрирован препарат конфидор из группы хлорникотинолов с дозировкой 10-15 г действующего вещества на гектар.
Значительно изменялись и способы распределения химических препаратов. Если в начале XX века использовали преимущественно очень примитивные и малопроизводительные ранцевые и конные опрыскиватели, то сейчас используются разнообразные технические средства, сочетание которых позволяет существенно уменьшить расход препарата, что важно как с экологической, так и с экономической точки зрения.
Для снижения численности кулиг весьма эффективны приманки, в качестве основы которых используют отруби или жмых. Но для приманок необходимо использовать инсектициды с длительным сроком действия (адонис, ингибиторы синтеза хитина).
Применение различных инсектицидов в виде дуста (т.е. в пылевой форме) также может быть оправданным, особенно при недостатке воды и оборудования.
Естественно, в настоящее время наиболее широко используются разного рода опрыскиватели. Традиционно используемые в России и других странах СНГ формы препаратов предназначены для разведения в больших объемах воды, что ограничивает их применение. В мире сейчас широко применяются разнообразные ультрамалообъемные опрыскиватели (УМО), распыляющие концентрированные растворы инсектицидов в масле. Использование этой технологии позволяет в целом значительно снизить расход препарата, в том числе за счет более мелкого распыления и (в современных конструкциях) за счет частично реализованной возможности регулирования размеров капель, а также почти избавляет от привязанности к источникам воды. Использование УМО дает возможность достаточно точно обрабатывать отдельные кулиги (в первую очередь с помощью ручных опрыскивателей либо опрыскивателей на наземном транспорте). Весьма удобна эта технология для обработки небольших площадок и создания барьеров.
Существуют конструкции, приспособленные для самых разных условий - от ручных до производительных автомобильных. В последние годы в мире именно они широко используются на легких самолетах и вертолетах, а также на мотодельтапланах. Это очень экономичные устройства, способные обработать значительные поверхности. Так как на них установлены приемники системы спутникового определения координат (GPS), это позволяет пилоту очень точно выйти на заданную цель. При этом достигается очевидная экономия, в том числе трудозатрат, связанных с обозначением участков, требующих обработки.
В странах СНГ (особенно в Казахстане и России) широко применяются аэрозольные генераторы различных конструкций. Использование предыдущего поколения таких установок против кулиг итальянского пруса показало его высокую эффективность (до 90%) [Применение ..., 1978]. Специальные эксперименты, проведенные в Северном Казахстане, Таджикистане и на юге Новосибирской области (но везде при сравнительно невысокой плотности саранчовых), показали возможность эффективного использования аэрозольных генераторов с регулируемой дисперсностью капель (ГРД) для управления численностью нестадных саранчовых и итальянского пруса [Соболев, Сергеев, 1985; Сергеев и др., 1988]. Самое существенное, что с помощью таких обработок можно относительно быстро и дешево снять пик численности и (с экологической точки зрения) привести популяции в “нормальное” состояние. В начале 90-х годов ГРД и его компрессорный вариант применялся в Центральной Якутии для снижения численности нестадных саранчовых. В 1997-1999 гг. - в Северном Казахстане для нестадных саранчовых и итальянского пруса. Летом 1999 и 2000 гг. ГРД привлекались к борьбе с прусом на юго-западе Новосибирской области. Достоинство этой технологии - в значительном уменьшении доз распределяемого препарата и в возможности выбора оптимального для объекта обработки размера капель. Однако большое облако мельчайших капель фактически неуправляемо. Поэтому для успешного использования этой техники против саранчовых необходимы специальные исследования, направленные на решение технологических проблем.
Значительное своеобразие может быть отмечено и при использовании инсектицидов против нестадных саранчовых:
(1) Частая необходимость неоднократных обработок против многовидовых комплексов. Если в Центральной Якутии такие доминанты, как сибирская и белополосая кобылки, развиваются практически одновременно, то на юге Сибири, Урала, в Предкавказье и Казахстане фенологические расхождения их популяций достигают 3-4 недель. А так как обработки наиболее целесообразны против личинок 2-3-го возрастов, то проблема становится очевидной даже при использовании препаратов с длительным сроком действия.
(2) Настоятельно необходим учет мозаичного использования земель во многих районах, например в лесостепях Сибири, горных регионах, а также в Центральной Якутии.
(3) Все это подчеркивает необходимость сокращения доз инсектицидов и целесообразность использования против нестадных саранчовых очень гибкого сочетания разных тактик обработок. В каких-то случаях оптимальны приманки. В других - использование ручных ультрамалообъемных опрыскивателей. В третьих - самолетные обработки, в том числе барьерные. Вместе с тем в современных условиях экономические условия и экологические требования часто не позволяют использовать традиционные сплошные авиаобработки.
К сожалению, в случае возникновения реальных чрезвычайных ситуаций без химических препаратов обойтись трудно, но именно поэтому очень важна разработка экологически оправданных предупредительных технологий и системы прогнозирования.
(3) Биологические подходы основаны на использовании других живых организмов для управления популяциями саранчовых. Нередко подобные методы контроля весьма специфичны, и поэтому круг нежелательных последствий много уже. Вместе с тем использование биологических объектов требует тщательных предварительных исследований, так как их внедрение (особенно в случае завоза микроорганизмов из других стран) может привести к непредсказуемым изменениям в экосистемах.
Для биологического контроля возможно использование нескольких групп объектов [Daxl et al., 1994]:
Сейчас нередко отдельно рассматривают биотехнологические методы [Daxl et al., 1994], к которым относят применение специально разводимых микроорганизмов и вирусов. Кроме того, в эту группу можно включить использование аттрактантов и репеллентов, регуляторов роста и их аналогов, в первую очередь получаемых из растений, а также генно-инженерными путями. Почти все эти методы по разным причинам мало применимы для саранчовых.
Однако в последнее десятилетие активно обсуждается возможность широкого использования азадирахтина - продукта переработки плодов тропических деревьев ним (Azadirachta indica и Melia volkensii из семейства Meliaceae) [Rembold, 1994 и др.]. Также вероятно и успешное применение веществ, используемых насекомыми для распознавания друг друга. С их помощью можно научиться управлять образованием кулиг и стай, а также их перемещением.
Есть перспективы обнаружения поражающих именно саранчовых форм одной из наиболее широко применяемых в биоконтроле бактерий - Bacillus thuringiensis, а также некоторых групп вирусов.
Яйца в кубышках часто выедаются личинками жуков-нарывников из родов Mylabris и Epicauta, а также личинками жужелиц из рода Amara и мух-жужжал родов Anastoechus и Cythrea [Мищенко, 1952]. Вероятно и поражение яиц специализированными яйцеедами, клещиками и различными грибами [Сергеев, Копанева, Рубцов и др., 1995].
Массовую гибель личинок и взрослых особей саранчовых может вызывать гриб Entomophaga grylli [Мищенко, 1952 и др.]. В экспериментах показана высокая эффективность грибов из рода Aspergillus (A. flavus, A. ochraceus), Beauveria sp. и микроспоридий Nosema sp. [Нуржанов, Лачининский, 1987].
В последние годы в тропических и субтропических районах для регулирования численности саранчовых довольно широко используются культуры грибов из рода Metarhizium. К сожалению, по сравнению с большинством химических препаратов они действуют медленно и наибольший эффект достигается лишь через несколько недель после обработки. Кроме того, грибы, поражающие насекомых, хорошо развиваются во влажные годы, тогда как в степях подъемы численности саранчовых приходятся на засухи. Зато такие грибы практически безопасны для подавляющего большинства остальных животных, в том числе человека (хотя, очевидно, могут выступать как аллергены).
Среди паразитов личинок и имаго хорошо представлены и насекомые, в частности мухи - Anastoechus nitidulus, Systoechus sp., Cytherea setosa, Blaesoxipha lineata, Wohlfartia balassogloi, а также клещики и мермитиды [Мищенко, 1952; Uvarov, 1977 и др.].
Естественна и гибель пруса при поедании хищниками. Это многочисленные членистоногие, ящерицы и птицы. Один из наиболее известных врагов пруса розовый скворец, огромные стаи которого кочуют в местах массового размножения саранчовых. Показано также, что грачи и испанские воробьи используют в пищу значительные количества саранчовых [Анохина, 1981].
(4) Агротехнические приемы (главным образом уничтожения кубышек) широко использовались и используются для регулирования численности саранчовых. Еще в 30-е гг. Лепешкин [1934в] рекомендовал распашку благоприятных для пруса (особенно для яйцекладки) местообитаний, их полив, искуственное загущение растительного покрова. В общем, распашка в разных вариантах приводит к уничтожению всех либо большинства кубышек. Вместе с тем это создает благоприятные условия для размножения пруса на следующий год.
Из агротехнические методов также может использоваться улучшение пастбищных и сенокосных угодий за счет подсева трав и соответствующего загущения растительного покрова и(или) за счет регулирования выпаса скота [Цыпленков, 1970]. Возможно использование растений, привлекательных для насекомых [Daxl et al., 1994], которые концентрируются вблизи или на них. Для управления популяциями итальянского пруса Л.Д. Бунин [1979] предлагал использовать определенную систему размещения полей различных культур, в том числе сосредоточение в районах регулярных подъемов численности этого вида посевов проса - злака, почти не повреждаемого им. Наилучший же вариант - ускорение восстановления степных экосистем на выведенных из хозяйственного оборота землях.
Современные подходы к управлению численностью саранчовых направлены на решение (хотя бы частичное) экологических и экономических проблем. Очевидно, что пока во время вспышек массового размножения без использования химических препаратов не обойтись. Поэтому в первую очередь, нужно четко представлять какие препараты, в каких ситуациях и в каких количествах предпочтительно использовать, какие технологии целесообразно применять в каждой конкретной ситуации.
Несомненно, необходимо стремиться к уменьшению площадей, обрабатываемых против саранчовых, и к сокращению вносимых доз. Это реально, если использовать барьерные и блочные обработки, когда препарат распространяется только на части защищаемой территории, например полосами, между которыми остаются необработанные промежутки. Часто достаточно создать преграду в виде полосы, препятствующей распространению личинок на поля.
Для активно перемещающихся саранчовых, например, пустынной саранчи, ширина межбарьерных участков может достигать 2 км. Экспериментально показано, что для итальянского пруса ширина необработанных полос должна быть существенно меньше - 100-300 м, а при низкой подвижности (например, при достаточном количестве кормовых растений, особенно для личинок младших и средних возрастов) их ширина вообще должна быть такой же, как для нестадных саранчовых, т.е. 15-40 м [см. “Рекомендации …”, 2000]. Наши эксперименты показали, что при избытке корма кулиги таких личинок вообще почти не перемещаются и тогда барьерные обработки мало эффективны.
Для барьерных и блочных обработок, а также для отравленных приманок, конечно, пригодны устойчивые препараты, например адонис и ингибиторы синтеза хитина. В то же время для быстрой защиты посевов оптимальны пиретроиды, фосфорорганика и адонис [Лачининский, 2000].
Барьерные обработки являются неотъемлемой частью недавно разработанного американскими специалистами интегрированного метода борьбы с нестадными саранчовыми - СИМП (Сниженный Инсектицид на Меньшей Площади - в английском варианте - RAATs) [Lockwood, Schell, 1997; Локвуд, Лачининский, Сергеев, 2000]. Эта стратегия основана на объединении химического и биологического методов управления численностью саранчовых. Используется меньшее количество инсектицида (т.е. уменьшается доза) и не обрабатывается весь защищаемый участок. Фактически сохраняются убежища для организмов менее подвижных, чем саранчовые. Благодаря этому хищники и паразиты, перемещающиеся с необработанных полос, способствуют подавлению численности саранчовых.
Если использовать этот подход, то количество инсектицида уменьшается на 50% и более. Кроме того, существенно сокращаются затраты на обработку. Конечно, достигаемая эффективность меньше (80-95% [Локвуд и др., 2000]), чем при сплошных обработках, но примечательно, что оставшиеся разреженные саранчовые не дают вспышек на следующий год. Это также важно, так как сохраняются популяции их паразитов и хищников.
Оптимизация распределения капель также важна для снижения количества вносимых препаратов. Специальные эксперименты, проводившиеся в Институте химической кинетики и горения СО РАН (Новосибирск), показали, что на насекомых разных размеров и с разным характером расположения волосков оседают капли различного диаметра. Поэтому за счет подбора диаметра аэрозольных капель можно достичь максимальной эффективности.
Другое перспективное направление - это создание биопрепаратов на базе местных возбудителей болезней, в первую очередь грибов.
Кроме того, давно известно, что оптимальны обработки против личинок младших и средних возрастов, когда необходимые дозы и площади обработок невелики. В это время возможно эффективное использование приманок и ранцевых ультрамалообьемных опрыскивателей, а также барьерных и краевых обработок. Вероятно, перспективно и применение биопрепаратов, в том числе грибных. Существенно и то, что на этих возрастах велика естественная смертность и ограничены перемещения [Соболев, Сергеев, 1985]. Кроме того, по данным Курдюкова и Наумовича [1984], личинки 4-го возраста наиболее устойчивы к инсектицидам. Для условий Каракалпакии также рекомендуются обработки против личинок 2-3-го возрастов [Нуржанов, Шамуратов, 1988].
Вместе с тем, несомненно, требуется решение проблемы на экологическом уровне. Во-первых, необходима организация системы долговременного слежения за населением саранчовых в степных ландшафтах с учетом разнообразия видов и популяций. Во-вторых, необходима минимизация воздействия человека на степные ландшафты, в частности регулирование выпаса и разработка мер по ускорению восстановления степных сообществ на залежных землях. В связи с этим целесообразно определение оптимального соотношения экосистем разных типов для каждого региона и соответственное изменение, например, площадей пастбищ, полей и целинных и (или) восстановленных степных экосистем. Необходимо снижение выпаса до уровня, характерного для природных степей с дикими копытными, а также такой подбор животных, чтобы воздействие на растения и почву было разнообразным. Крайне желательно развитие особых технологий по восстановлению степных экосистем на брошенных землях, в том числе находящихся на крайних стадиях деградации после сбоя или многолетнего использования под поля однолетников. Это особенно актуально для районов, где почти не сохранились фрагменты степных экосистем и их производных, которые можно использовать в качестве своеобразных резерватов для восстановления.
Мы недооцениваем саранчовых и не уделяем им достаточное внимание. Со стратегической точки зрения необходимо осуществление следующих мероприятий:
(1) Тщательное эколого-географическое изучение популяционной динамики всех видов, стадных и нестадных. Это насущная потребность.
(2) Ориентация не на подавление вспышки саранчовых, а на ее предупреждение, то есть на то, что Н. Джэго [Jago, 1990] назвал превентивной стратегией контроля.
(3) Изменение самих сельскохозяйственных технологий: рациональное с эколого-географической точки зрения размещение полей, снятие перевыпаса. Ускорение процессов восстановления степных экосистем на залежных землях. Однако новые проблемы могут возникнуть в результате становления частного землевладения.
(4) Разработка самостоятельной стратегии управления численностью нестадных саранчовых с учетом особенностей умеренных экосистем. Такая стратегия должна, в частности, включать связанные друг с другом, но разнообразные тактические приемы, ориентированные на решение проблем конкретных районов, местообитаний и временных промежутков, а также учитывающих экономическую ситуацию.
Длительные исследования в области динамики и зонально-ландшафтного распределения популяций массовых видов, а также многовидовых сообществ саранчовых позволят выделить те критические местообитания и временные промежутки, которые существенны для динамики саранчовых. Такие данные могут служить основой для обоснования слежения, а также мер по управлению популяциями, ориентированных на минимизацию экологического ущерба и поддержание биоразнообразия [Sergeev, Denisova, Vanjkova, 2000].
Крайне актуально создание межгосударственных структур по управлению численностью саранчовых, так как многие районы, в пределах которых она может резко нарастать, располагаются в приграничных районах: в первую очередь вдоль почти всей границы России и Казахстана, а также в пограничных районах Казахстана, Киргизии, Узбекистана, Таджикистана и Китая.
Все это должно позволить не бросать деньги на ветер, не заниматься экстренным тушением пожара, а целенаправленно и целесообразно управлять населением саранчовых как компонентом степных экосистем.
Без саранчовых, судя по имеющимся данным, степные экосистемы не могут нормально функционировать. Эти насекомые - важнейшее звено круговорота вещества и энергии в степях, и в случае их исчезновения этот круговорот окажется нарушенным или даже полностью разорванным. Поэтому, если мы хотим сохранить степи (даже исходя из необходимости решения таких “корыстных” задач, как обеспечение продовольствием), мы должны сохранить и разнообразие саранчовых, деятельность которых ускоряет круговорот вещества и энергии и способствует сохранению устойчивости на ландшафтном уровне. Хотя мировой опыт показывает, что человек способен полностью уничтожить даже стадные виды саранчовых (например, так называемую саранчу Скалистых гор - серьезнейшего вредителя в североамериканских прериях XIX века [Lockwood, DeBrey, 1990]), тем не менее, в подавляющем большинстве случаев деятельность человека в степных ландшафтах - перевыпас скота, увеличение залежных земель, создание придорожных полос - способствует формированию вспышек массового размножения. Таким образом, создается частично замкнутый круг, который необходимо разорвать. А сделать это можно сделать не только путем разработки изощренных методов воздействия на популяции саранчовых, но и путем оптимизации отношения саранчовых и человека.
Алейникова М.М. Азиатская саранча в Татарской АССР // Изв. Каз. фил. АН СССР, сер. биол. и сельскохоз. наук. 1950. №2. С. 209-258.
Александров Ю.Д. Система землепользования и фауна вредных насекомых в Алтайском крае // Проблемы зоологических исследований в Сибири. Горно-Алтайск, 1962. С. 9-11.
Анохина Ю.Р. О роли гнездовых колоний грачей и испанских воробьев в снижении численности прямокрылых // Вопросы экологии. Новосибирск: изд. НГУ, 1981. С. 85-96.
Архангельский П.П. Саранчовые в Казахстане подавлены // Защита растений от вредителей и болезней. 1957. №6. С. 17-20.
Бей-Биенко Г.Я. Отряд Orthoptera (Saltatoria) - Прямокрылые (прыгающие прямокрылые) // Определитель насекомых европейской части СССР. Т. 1. М., Л.: Наука, 1964. С. 205-284.
Бей-Биенко Г.Я., Мищенко Л.Л. Саранчовые фауны СССР и сопредельных стран. Т. 1-2. М., Л.: Изд-во АН СССР, 1951. 667 с.
Бережков Р.П. Саранчовые Западной Сибири. Томск: Изд-во ТГУ, 1956. 175 с.
Борисенков Е.П., Пасецкий В.М. Тясячелетняя летопись необычайных явлений природы. М.: Мысль, 1988. 524 с.
Бугаев Г.С. Стациальное распределение нестадных саранчовых в зоне ленточных сосновых боров в Северо-Восточном Казахстане // Вестн. с.-х. науки Казахстана. 1977. №6. С. 37-40.
Бунин Л.Д. Итальянская саранча (Calliptamus italicus L.)в зоне почвозащитного земледелия на востоке Казахстана и совершенствование мероприятий по борьбе с ней: Автореф. канд. дис. Л., 1979. 24 с.
Ванькова И.А. Опыт изучения фенотипической изменчивости саранчовых рода Calliptamus Serv. (Orthoptera, Acrididae) // Биологическое разнообразие животных Сибири, Томск, 1998. С. 35-36.
Васильев К.А. Миграционные перелеты у итальянской саранчи (Calliptamus italicus L.) // Докл. АН СССР. 1950а. Т. 74, №2. С. 385-388.
Васильев К.А. Фазы у итальянской саранчи (Calliptamus italicus L.) // Докл. АН СССР. 1950б. Т. 74, №3. С. 639-642.
Васильев К.А. Итальянская саранча (Calliptamus italicus L.) в Центральном Казахстане // Тр. НИИ защиты растений КазССР. 1962. Т. 7. С. 124-190.
Васильев К.А. Вредные саранчовые в зоне освоения целинных и залежных земель // Тр. Всесоюз. энтомол. о-ва. 1965. Т. 50. С. 129-145.
Волков А.Н. Борьба с азиатской саранчой. М.: Сельхозгиз, 1947. 80 с.
Гусева В.С., Крицкая И.Г. Изменения в фауне саранчовых за последние полвека в заповедниках степной зоны европейской части СССР // Уч. зап. МГПИ. 1970. №394. С. 64-73.
Гусева В.С., Литвинова Н.Ф., Черняховский М.Е. Определение вредных саранчовых и меры борьбы с ними. М.: изд. ЦНИТ Министерства сельского хозяйства и продовольствия РФ, 1994. 10 с.
Зряковский В.Н. Азиатская саранча и меры борьбы с нею. Пятигорск: изд. Терского окружного совета ОСОАвиахима, 1927. 22 с.
Камбулин В.Е., Бугаев Г.С. Значение нестадных саранчовых (Orthoptera, Acrididae) в злаковых ассоциациях западной части Центрального Казахстана и зоны ленточных боров Северо-Восточного Казахстана // Энтомол. обозр. 1980. Т. 59, №3. С. 529-534.
Кобахидзе Д.Н., Абашидзе Э.Д. К изучению качественно-количественных изменений акридофауны в связи с сельскохозяйственным освоением земель на примере Самгорской степи (Грузинская ССР) // Тр. Ин-та защиты растений ГрузССР. 1970. Т. 22. С. 61-63.
Копанева Л.М., Надворный В.Г., Стебаев И.В. Распределение прямокрылых в долинах рек в связи с комплексным подходом к охране энтомофауны и защите растений на примере Днепра и Иртыша // Исследования по энтомологии и акарологии на Украине. Тез. докл. 2-го Съезда УЭО. Киев, 1980. С. 36-38.
Курдюков В.В., Васильев С.В. Обоснование разработки экспресс-метода определения эффективности применения химических средств борьбы с саранчовыми // Саранчовые -- экология и меры борьбы. Л.: изд. ВИЗР, 1987. С. 9298.
Курдюков В.В., Наумович О.Н. Видовая и межвидовая изменчивость чувствительности саранчовых к инсектицидам // Бюл. ВНИИ защиты растений. 1984. №58. С. 7-12.
Куркин К.А. Системные исследования динамики лугов. М.: Наука, 1976. 284 с.
Лачининский А.В. Новые препараты для борьбы с вредными саранчовыми // Защита и карантин растений. 2000. №4. С. 9-11.
Лачининский А.В., Локвуд Дж.А., Сергеев М.Г. Опыт борьбы с саранчовыми североамериканских прерий // Защита и карантин растений. 1999. №8. С. 12-14.
Лепешкин С.Н. О ликвидации пруса (Calliptamus italicus L.) в Мервском оазисе // Саранчовые Средней Азии. М., Ташкент, 1934а. С. 9-81.
Лепешкин С.Н. Регистрация пруса в поливных районах Средней Азии // Саранчовые Средней Азии. М., Ташкент, 1934б. С. 237-243.
Лепешкин С.Н. Предварительная система мероприятий по борьбе с оазисным прусом // Саранчовые Средней Азии. М., Ташкент, 1934в. С. 229-236.
Лер П.А. Материалы по биологии туранского и пустынного прусов (Orthoptera, Calliptamus) в Чимкентской области // Тр. КазИЗР. 1962. Т. 7. С. 3-56.
Литвинова Н.Ф., Гусева В.С., Воронцова Л.И. Трансформация комплексов саранчовых при антропогенном воздействии в условиях сухих степей Нижнего Поволжья // Зоол. журн. 1994. Т. 73, № 10. С. 16-25.
Локвуд Дж.А., Бомар Ч.Р., Уильямс С.Е., Додд Дж.Л., Цуан М., Ли Х. Экология насекомых в азиатских и североамериканских степях: поражающие различия и замечательное сходство // Сибирский экол. журн. 1997. Т. 4, №3. С. 241-251.
Локвуд Дж.А., Лачининский А.В., Сергеев М.Г. Новая стратегия обработок против нестадных саранчовых на пастбищах // Защита и карантин растений. 2000. №7. С.10-11.
Махоткин А.Г., Зверев А.А., Махоткина Л.Я. Обработки почвы против пруса // Защита и карантин растений. 1999. №8. С. 19.
Мищенко Л.Л. Саранчовые (Catantopinae). М., Л.: Изд-во АН СССР, 1952. 610 с. (Фауна СССР. Насекомые прямокрылые. Т. 4, вып. 2).
Насырова С.Р. Влияние сельскохозяйственного освоения степей Прииртышского плато на распределение и численность саранчовых (Orthoptera, Acrididae) // Тр. Ин-та зоологии АН КазССР. 1990. Т. 45. С. 93-103.
Наумович О.Н., Павлюченко А.А. Видовой состав и рапространение
короткоусых прямокрылых (Orthoptera, Caelifera) Внутреннего Тянь-Шаня // Саранчовые -- экология и меры борьбы. Л.: изд. ВИЗР, 1987. С. 38-51.
Нуржанов А.А., Лачининский А.В. Энтомопатогенные микроорганизмы стадных саранчовых в Узбекистане // Саранчовые -- экология и меры борьбы. Л.: изд. ВИЗР, 1987. С. 62-69.
Нуржанов А.А., Шамуратов Г.Ш. Итальянский прус в Каракалпакии и химический метод борьбы с ним // Защита сельскохозяйственных культур от основных вредителей и сорняков в Каракалпакской АССР. Нукус, 1988. С. 98-107.
Определитель сельскохозяйственных вредителей по повреждениям
культурных растений. Л.: Колос, 1976. 696 с.
Применение аэрозолей для борьбы с вредными насекомыми. Новосибирск: Наука, 1978. 150 с.
Пшеницына Л.Б. Пищевая избирательность саранчовых в связи с их воздействием на степные фитоценозы: Автореф. канд. дис. Новосибирск: Биологический ин-т СО АН СССР, 1987. 18 с.
Раевский В.Г. Наблюдения за прусом (Calliptamus italicus L.) в Славгородском уезде в 1922 году // Изв. Сиб. энтомол. бюро. 1924. Т. 2. С. 53-58.
Рекомендации по мониторингу и борьбе с вредными саранчовыми. М.: Колос, 2000. 21 с.
Рубцов И.А. О количестве пищи, поедаемой саранчовыми // Защита растений. 1932. №2. С. 31-40.
Семевский Ф.Н. Об оценке эффективности борьбы с насекомыми // Зоол. журн. 1968. Т. 47, №8. С. 1233-1236.
Сергеев М.Г. Секторная дифференциация населения прямокрылых насекомых лесостепной, степной и полупустынной зон // Изв. СО РАН, сер. биол. наук. 1990. №3. С. 85-89.
Сергеев М.Г. Закономерности распределения насекомых-фитофагов в травянистых экосистемах Голарктики // Изв. АН. Сер. биол. 1998. №4. С. 445-450.
Сергеев М.Г., Бугров А.Г., Казакова И.Г., Соболев Н.Н. Регуляция динамики популяций саранчовых в агроландшафтах с помощью инсектицидных аэрозолей // Ландшафтная экология насекомых. Новосибирск: Наука, 1988. С. 63-69.
Сергеев М.Г., Ванькова И.А. Зонально-ландшафтное распределение
популяционных группирок итальянского пруса Calliptamus italicus L. (Insecta, Orthoptera, Acrididae) // Сиб. экол. журн. 1996. Т. 3, №3-4. С. 219-225.
Сергеев М.Г., Копанева Л.М., Рубцов И.А., Антипанова Е.М., Бугров А.Г., Высоцкая Л.В., Иванова И.В., Казакова И.Г., Карелина Р.И., Пшеницына Л.Б., Соболев Н.Н., Чогсомжав Л. Сибирская кобылка (Aeropus sibiricus L.). Новосибирск: Наука, 1995. 176 с.
Соболев Н.Н., Сергеев М.Г. Популяционная динамика саранчовых (Orthoptera, Acrididae) в агроценозах Северного Казахстана // Антропогенные воздействия на сообщества насекомых. Новосибирск: Наука, 1985. С. 96-104.
Стебаев И.В. Характеристика надпочвенного и напочвенного зоомикробиологических комплексов степных ландшафтов Западной и Средней Сибири // Зоол. журн. 1968. Т. 47, №5. С. 661-675.
Столяров М.В. Итальянская саранча (Calliptamus italicus L.) в Западном Казахстане // Тр. Всесоюз. энтомол. об-ва. 1974. Т. 57. С. 98-111.
Стороженко С.Ю. Отряд Orthoptera (Saltatoria) - Прямокрылые (прыгающие прямокрылые) // Определитель насекомых Дальнего Востока СССР. Т. 1. Л.: Наука, 1986. С. 241-317.
Цыпленков Е.П. Вредные саранчовые насекомые в СССР. Л.: Колос, 1970. 271 с.
Четыркина И.А. Распространение и зоны вредоносности пруса (Calliptamus italicus L.) в Казахстане // Итоги научно-исследовательских работ ВИЗР за 1935 г. Л., 1936. С. 20-22.
Четыркина И.А. Саранчовые лесных опушек в долине р. Урала // Тр. Зоол. ин-та АН СССР. 1952. Т. 11. С. 133-141.
Четыркина И.А. Прус или итальянская саранча Calliptamus italicus L. в Восточном Казахстане // Тр. Всесоюз. энтомол. об-ва. 1958. Т. 46. С. 5-67.
Щербиновский Н.С, Пустынная саранча шистоцерка. М.: Сельхозгиз, 1952. 416 с.
Daxl R., von Kayserlingk N., Klein-Koch C., Link R., Waibel H. Integrated pest management. Rossdorf: TZ-Verl.-Gez., 1994. 119 p.
P. 1-25.
Harz K. Die Tierwelt Deutschlands und der angrenzenden Meersteile. 46. Teil. Gerdflügler oder Orthopteren (Blattodea, Mantodea, Saltatoria, Dermaptera). Jena: Gustav Fischer Verlag, 1960. 232 S.
Jago N.D. A revision of the genus Calliptamus Serville (Orthoptera: Acrididae) // Bull. Brit. Mus. (Nat. Hist.) Ent. 1963. Vol. 13, N 9. P. 287-350.
Jago N.D. The present and future roles of the Orthopterist // Bol. San. Veg. Plagas (Fuera de serie). 1990. Vol. 20. P. 1-8.
Lockwood J.A., DeBrey L.D. A solution for the sudden and unexplained extinction of the Rocky Mountain grasshopper (Orthoptera: Acrididae) // Environ. Entomol. 1990. Vol. 19. P. 1194-1205.
Lockwood J.A., Schell S.P. Decreasing economic and environmental costs through Reduced Area and Agent Insecticide Treatments (RAATs) for the control of rangeland grasshoppers: empirical results and their implications for pest management // J. Orthoptera Research. 1997. N 6. P. 19-32.
Louveaux A., Peyrelongue J.-Y., Gillon Y. Analyse des facteurs de pullulation du criquet italien (Calliptamus italicus L.) en poitou-charentes // C. r. Acad. agr. Fr. 1988. T. 74, N 8. P. 91-102.
Olfert O.O., Mukerji M.K. Effects of acute simulated and acute grasshopper (Orthoptera: Acrididae) damage on growth rates and yield of spring wheat (Triticum aestivum) // Can. Ent. 1983. Vol. 115, N 6. P. 629-639.
PRIFAS. La lutte preventive anti-SGR // Surveillance des acridiens au Sahel. 1989. N 6. P. 28.
Rembold H. Controlling locusts with plant chemicals // New trends in locust control. Rossdor: TZ-Verl.-Ges., 1994. P. 41-49.
Ritchie J.M., Dobson H. Desert locust control operations and their environmental impacts // NRI Bulletin. 1995. Vol. 67. vi+42 p.
Sergeev M.G., Denisova O.V., Vanjkova I.A. How do spatial population structures affect grasshopper and locust management? // Grasshopper and grassland health. Dordrecht et al.: Kluwer Academic Publ., 2000. P. 71-87.
Uvarov B.P. Grasshoppers and locusts: A handbook of general acridology. Vol. 1 Cambridge: Univ. Press, 1966. 481 p.
Uvarov B.P. Grasshoppers and locusts: A handbook of general acridology. Vol. 2. London: Centre for Overseas Pest Research, 1977. 613 p.
Wewetzer A., Krall S., Schulz F.A. Methods for the assessment of crop losses due to grasshoppers and locusts. Rossdorf: TZ-Verl.-Gez., 1993. 54 p.
Аттрактант - вещество, привлекающее животных, особенно насекомых.
Возраст - здесь состояние личинки между промежуточными линьками; личинка 1-го возраста вылупляется из яйца; личинка последнего возраста у саранчовых линяет во взрослое насекомое.
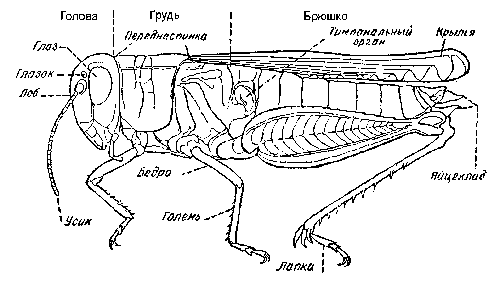
Грудной отдел - часть тела насекомых, включающая 3 сегмента, несущих 3 пары двигательных конечностей и у большинства взрослых насекомых 2 пары крыльев. Располагается между головным и брюшным отделами.
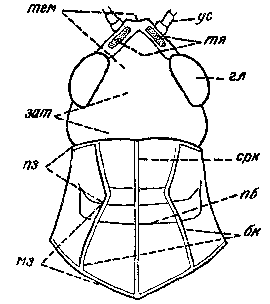
Заднеспинка - здесь верхняя часть заднего сегмента (последнего из трех) грудного отдела у насекомых.
Ингибитор - вещество, снижающее скорость химической реакции или вообще подавляющее ее.
Кубышка - у большинства саранчовых специальная структура образуемая во время яйцекладки: в ее состав обычно входят яйца, окружающие их специальные выделения и частицы почвы.
Кулига - стая личинок саранчовых, обычно плотное скопление особей, перемещающихся при необходимости в одном направлении.
Ландшафт - генетически целостная географическая система, неделимая по зональным и азональным признакам, с единым геологическим фундаментом, однотипным рельефом, общим климатом, единообразным сочетанием гидротермических условий, почв, растительности, животного населения и характерным устройством.
Личинка - стадия индивидуального развития насекомых; у саранчовых вылупляется из яйца, претерпевает несколько линек, переходя их одного возраста в другой, и, наконец, линяет во взрослое насекомое. Личинки не способны размножаться и не могут летать.
Мониторинг -- длительное слежение за состоянием какого-то объекта.
Надкрылья - у саранчовых уплотненная передняя пара крыльев, отходящая от среднего сегмента груди; в состоянии покоя сложены вдоль брюшка и прикрывают заднюю пару крыльев.
Переднеспинка - здесь верхняя часть переднего сегмента грудного отдела насекомых. Обычно очень хорошо развита и прикрывает сверху средне- и заднеспинку, а также основания крыльев.
Подсемейство - группа, объединяющая родственные трибы или роды (если трибы не выделяются); близкие подсемейства относятся к одному семейству.
Популяция - совокупность особей одного вида, занимающая на протяжении ряда поколений один и тот же участок земной поверхности и более или менее изолированная от соседних популяций того же вида.
Превентивный - предваряющий, предупреждающий.
Репеллент - препарат, отпугивающий животных.
Саранча - обычно используется для обозначения стадных саранчовых.
Саранчовые - обычно все представители короткоусых прямокрылых насекомых, кроме триперстовых и прыгунчиков.
Сегмент - он же членик, часть тела насекомого, снаружи обычно представленная 2 полукольцами (верхним и нижним); сегменты идут один за другим; в состав грудного отдела входит 3 сегмента, а брюшного - 11 (но обычно не все из них различимы).
Среднеспинка - здесь верхняя часть среднего сегмента грудного отдела насекомых.
Тимпанальный орган - здесь орган слуха у саранчовых, расположенный по бокам 1-го сегмента брюшка. Обычно хорошо развит и представлен овальным отверстием с мембраной.
Трансекта - узкая и длинная площадка или линия, пересекающая соседние участки (разные места обитания); в ее пределах или вдоль нее ведут учеты численности.
Триба - группа родственных родов; обычно несколько триб объединяют в одно подсемейство.
Хитин - очень прочный сложный полисахарид с участием азотных групп. У насекомых образует наружный скелет, у грибов - клеточную стенку.
Церки - придатки последних сегментов тела у многих насекомых; у саранчовых короткие нечленистые образования.
Экологическая ниша - характеризует место вида в экосистеме, в том числе его функциональную роль.
Экосистема - любое определенное во времени и пространстве сообщество живых существ и его среда обитания, объединенные в единое функциональное целое, возникающее на основе внутренних и внешних связей.


| © 2000-2026 гг. Центр охраны дикой природы. Все права защищены |